Классификация микроорганизмов по их основным формам. Бактерии, их разнообразие. Строение. Жизнедеятельность. Базовые термины: бактерии
Питание бактерий.
Питание.
Пассивная диффузия
Облегчённая диффузия
Активный транспорт
В первом случае молекула питательного вещества образует комплекс с белком периплазматического пространства, который взаимодействует со специфической пермеазой цитоплазматической мембраны. После энергозависимого проникновения через цитоплазматическую мембрану комплекс «субстрат – белок периплазмы – пермеаза» диссоциирует с освобождением молекулы субстрата.
При активном транспорте с химической модификацией переносимого вещества цепь событий включает: (1) фосфорилирование мембранного фермента-2 со стороны цитоплазмы фосфоенолпируватом; (2) связывание на поверхности цитоплазматической мембраны фосфорилированным ферментом-2 молекулы субстрата; (3) энергозависимый транспорт молекулы субстрата в цитоплазму; (4) перенос фосфатной группы на молекулу субстрата; (5) диссоциация комплекса «субстрат – фермент» в цитоплазме. За счёт фосфорилирования молекулы субстрата аккумулируются в цитоплазме клеток и не способны выйти из них.
Классификация бактерий по типу питания.
По способу поступления питательных веществ бактерии подразделяются на голофиты и голозои . Бактерии-голофиты (от греч. holos – полноценный и phyticos – относящийся к растениям) неспособны выделять в окружающую среду ферменты, расщепляющие субстраты, вследствие чего потребляют питательные вещества исключительно в растворённом, молекулярном виде. Бактерии-голозои (от греч. holos – полноценный и zoikos – относящийся к животным), напротив, имеют комплекс экзоферментов, которые обеспечивают внешнее питание – расщепление субстратов до молекул вне бактериальной клетки. После этого молекулы питательных веществ поступают внутрь бактерий-голозоев.
По источнику углерода среди бактерий выделяют автотрофы и гетеротрофы . Автотрофы (от греч. autos – сам, trophe – пища) в качестве источника углерода используют углекислый газ (СО 2), из которого синтезируют все углеродсодержащие вещества. Для гетеротрофов (от греч. geteros – другой и trophe – пища) источником углерода являются различные органические вещества в молекулярной форме (углеводы, многоатомные спирты, аминокислоты, жирные кислоты). Наибольшая степень гетеротрофности присуща прокариотам, которые могут жить только внутри других живых клеток (например, риккетсии и хламидии).
По источнику азота прокариоты подразделяются на 3 группы: 1) азотфиксирующие бактерии (усваивают молекулярный азот из атмосферного воздуха); 2) бактерии, потребляющие неорганический азот из солей аммония, нитритов или нитратов; 3) бактерии, которые ассимилируют азот, содержащийся в органических соединениях (аминокислоты, пурины, пиримидины и др.).
По источнику энергии бактерии делят на фототрофы и хемотрофы . Бактерии-фототрофы , как и растения, способны использовать солнечную энергию. Фототрофные прокариоты заболеваний у человека не вызывают. Бактерии-хемотрофы получают энергию при окислительно-восстановительных реакциях.
По природе доноров электронов литотрофы (от греч. litos – камень) и органотрофы . У литотрофов (хемолитотрофов ) в качестве доноров электронов выступают неорганические вещества (Н 2 , Н 2 S, NH 3 , сера, CO, Fe 2+ и др.). Донорами электронов у органотрофов (хемоорганотрофов ) являются органические соединения – углеводы, аминокислоты и др.
Большинство патогенных для человека бактерий обладает хемоорганотрофным (хемогетеротрофным) типом питания; реже встречается хемолитотрофный (хемоавтотрофный) тип.
По способности синтезировать органические соединения бактерии-хемотрофы подразделяются на прототрофы, ауксотрофы и гипотрофы . Бактерии-прототрофы синтезируют из глюкозы и солей аммония все необходимые органические вещества. Бактерии называются ауксотрофами , если они неспособны синтезировать какое-либо органическое вещество из указанных соединений. Крайняя степень утраты метаболической активности называется гипотрофией. Гипотрофные бактерии обеспечивают свою жизнедеятельность, реорганизуя структуры или метаболиты хозяина.
Кроме углерода и азота, для полноценной жизнедеятельности бактериям необходимы сера, фосфор, ионы металлов. Источниками серы являются аминокислоты (цистеин, метионин), витамины, кофакторы (биотин, липоевая кислота и др.), сульфаты. Источниками фосфора служат нуклеиновые кислоты, фосфолипиды, фосфаты. В достаточно высоких концентрациях бактериям нужны магний, калий, кальций, железо; в значительно меньших – цинк, марганец, натрий, молибден, медь, никель, кобальт.
Факторы роста – это вещества, которые бактерии самостоятельно синтезировать не могут, но крайне в них нуждаются. В качестве факторов роста могут выступать аминокислоты, азотистые основания, витамины, жирные кислоты, железопорфирины и другие соединения. Для создания оптимальных условий жизнедеятельности бактерий факторы роста должны быть добавлены в питательные среды.
Метаболизм, превращение энергии
А) Конструктивный метаболизм.
Обязательной фазой питания бактерий является усвоение питательных веществ, то есть включение их в изменённом или модифицированном виде в синтетические реакции по воспроизведению клеточных компонентов и структур.
Белковый обмен у бактерий может протекать в 3 фазы: первичный распад белка, вторичный распад и синтез белка. Первичный распад белковых молекул до пептонов осуществляют экзоферменты – экзопротеазы, выделяемые бактериями в окружающую среду. Вторичный распад происходит под действием эндоферментов (эндопротеаз), которые имеют все бактерии. Этот процесс протекает внутри бактериальной клетки и заключается в расщеплении пептидов до составляющих их аминокислот. Последние могут быть использованы в неизменённом виде или быть подвергнуты химическим преобразованиям (дезаминирование, декарбоксилирование и др.), в результате которых появляются аммиак, индол, сероводород, кетокислоты, спирт, углекислый газ и др. Обнаружение указанных соединений имеет в бактериологии диагностическое значение.
Наряду с реакциями расщепления белков, происходят реакции их синтеза. Одни бактерии образуют белки из готовых аминокислот, полученных в результате внешнего питания, другие бактерии самостоятельно синтезируют аминокислоты из простых соединений, содержащих азот и углерод. Синтез аминокислот может осуществляться в реакциях аминирования, переаминирования, амидирования, карбоксилирования. Большинство прокариот способны синтезировать все аминокислоты, входящие в состав клеточных белков. Особенностью биосинтеза аминокислот является использование общих биосинтетических путей: цикл трикарбоновых кислот, гликолиз, окислительный пентозо-фосфатный путь и др. Основным исходным соединением для синтеза аминокислот является пируват и фумарат.
Углеводный обмен у автотрофов и гетеротрофов имеет отличия (схема 1). Бактерии-автотрофы все необходимые углеводы синтезируют из углекислого газа. Сырьём для образования углеводов у бактерий-гетеротрофов могут служить: (1) одно-, двух- и трёхуглеродные соединения; и (2) полисахариды (крахмал, гликоген, целлюлоза). Для расщепления последних многие бактерии-гетеротрофы имеют экзоферменты (амилаза, пектиназа и др.), которые проводят гидролиз полисахаридов до образования глюкозы, мальтозы, фруктозы и пр.
У бактерий-автотрофов в цикле Кальвина из углекислого газа образуется рибулозофосфат-фосфорно-глицериновая кислота, которая включается в реакции гликолиза, идущие в обратном направлении. Конечным продуктом обратного синтеза является глюкоза.
Бактерии-гетеротрофы образуют глюкозу из одно-, двух- и трёхуглеродных соединений, также включая их в реакции обратного гликолиза. Ввиду того, что некоторые реакции гликолиза необратимы, у гетеротрофов сформировались специальные ферментативные реакции, позволяющие обходить необратимые реакции катаболического пути.
При расщеплении бактериями-гетеротрофами полисахаридов образующиеся дисахариды поступают внутрь клеток и под влиянием мальтозы, сахарозы, лактозы подвергаются гидролизу и распаду на моносахара, которые затем сбраживаются либо включаются в реакции взаимопревращения сахаров.
Липидный обмен . Исходными материалами для образования липидов у бактерий могут служить как экзогенные липиды, так и амфиболиты межуточного обмена. Экзогенные липиды подвергаются действию бактериальных липаз и других липолитических ферментов. Многие виды бактерий усваивают глицерин, который служит источником пластического материала и энергии. Эндогенными источниками для синтеза липидов могут быть ацетилкоэнзим А, пропионил-АПБ, малонил-АПБ (АПБ – ацетилпереносящий белок), фосфодиоксиацетон и др.
Исходным субстратом для синтеза жирных кислот с чётным числом углеродных атомов служит ацетилкоэнзим А, для жирных кислот с нечётным числом углеродных атомов – пропионил-АПБ и малонил-АПБ. Образование двойных связей в молекуле кислоты у аэробных прокариот происходит при участии молекулярного кислорода и фермента десатуразы. У анаэробных прокариот двойные связи вводятся на ранних этапах синтеза в результате реакции дегидратации. Исходным субстратом для синтеза фосфолипидов служит фосфодиоксиацетон (промежуточное соединение гликолитического пути), восстановление которого приводит к образованию 3-фосфороглицерина. К последнему затем присоединяются 2 остатка жирных кислот в виде комплекса с АПБ. Продуктом реакции является фосфатидная кислота, активирование которой с помощью ЦТФ и последующее присоединение к фосфатной группе серина, инозита, глицерина или другого соединения приводят к синтезу соответствующих фосфолипидов.
Ауксотрофные и гипотрофные по жирным кислотам микроорганизмы (например, микоплазмы) получают их в готовом виде из клеток хозяина или питательной среды.
Мононуклеотидный обмен . Пуриновые и пиримидиновые мононуклеотиды являются важнейшими компонентами ДНК и РНК. Многие прокариоты способны как использовать содержащиеся в питательной среде готовые пуриновые и пиримидиновые основания, их нуклеозиды и нуклеотиды, так и синтезировать их из низкомолекулярных веществ. Бактерии располагают ферментами, катализирующими следующие этапы взаимопревращений экзогенных пуриновых и пиримидиновых производных: азотистое основание – нуклеозид – нуклеотид (моно- – ди- – трифосфат).
Синтез пуриновых и пиримидиновых мононуклеотидов de novo осуществляется независимыми путями. При синтезе пуриновых нуклеотидов в результате последовательных ферментативных реакций образуется инозиновая кислота, из которой путём химических модификаций пуринового кольца синтезируются адениловая (АМФ) и гуаниловая (ГМФ) кислоты. Синтез пиримидиновых нуклеотидов начинается с образования оротидиловой кислоты, декарбоксилирование которой даёт уридиловую кислоту (УМФ). Из последней образуется УТФ, ацилирование которого приводит к возникновению ЦТФ.
Дезоксирибонуклеотиды образуются в результате восстановления соответствующих рибонуклеотидов на уровне дифосфатов или трифосфатов. Синтез специфического для ДНК нуклеотида – тимидиловой кислоты происходит путём ферментативного метилирования дезоксиуридиловой кислоты.
Ионный обмен . Минеральные соединения – ионы, NH 3 + , К + , Mg 2+ , Fe 2+ , SO 4 2- , PO 4 3- и другие бактерии получают из окружающей среды как в свободном, так и в связанном с другими органическими веществами состоянии. Катионы и анионы транспортируются в бактериальную клетку различными способами, описанными в § 3. На скорость проникновения ионов в бактериальную клетку влияют рН среды и физиологическая активность самих микроорганизмов.
Б) Дыхание бактерий (энергетический метаболизм).
Все процессы жизнедеятельности энергозависимы, поэтому добывание энергии является крайне важной стороной метаболизма прокариот. Они получают энергию при анаэробном и аэробном дыхании.
Дыхание , или биологическое окисление – это катаболический процесс переноса электронов от вещества-донора на вещество-акцептор, сопровождающийся накоплением энергии в макроэргических соединениях . Дыхание осуществляется в процессе катаболических реакций, в результате которых сложные органические вещества, расщепляясь, отдают энергию и превращаются в простые соединения. Аккумулированная в макроэргических веществах (АТФ, ГТФ, УТФ и др.) энергия используется в анаболических реакциях.
По способу дыхания микроорганизмы подразделяются на облигатные (строгие) аэробы, облигатные анаэробы и факультативные анаэробы .
Облигатные аэробы нуждаются в свободном кислороде. Донорами электронов у патогенных для человека аэробов-хемоорганотрофов являются органические соединения (углеводы, жиры, белки), акцептором электронов – молекулярный кислород. Запасание энергии в виде АТФ у аэробов-хемоорганотрофов происходит при окислительном фосфорилировании доноров электронов. Аэробы обладают цитохромами (участвуют в переносе электронов), а также ферментами (каталаза, супероксиддисмутаза, пероксидаза), инактивирующими токсические кислородные радикалы, образующиеся при дыхании. Супероксиддисмутаза инактивирует наиболее токсичный метаболит – супероксидрадикал в Н 2 О 2 . Фермент каталаза превращает Н 2 О 2 в Н 2 О и О 2 .
Особую группу аэробов составляют микроаэрофильные бактерии , которые хотя и нуждаются в кислороде для получения энергии, лучше растут при повышенном содержании СО 2 , например, бактерии родов Campylobacter и Helicobacter .
Облигатные анаэробы не нуждаются в свободном кислороде, напротив, даже в малых количествах кислород оказывает на них токсическое действие. Донорами электронов у патогенных для человека анаэробов-хемоорганотрофов служат различные органические соединения (преимущественно углеводы). Акцептором электронов у анаэробов-хемоорганотрофов являются органические кислородсодержащие соединения – кислоты или кетоны, то есть акцептор электрона - связанный с органическим фрагментом кислород. Запасание энергии у этих прокариот происходит при субстратном фосфорилировании. Облигатные анаэробы, как правило, не имеют цитохромов и ферментов, инактивирующих кислородные радикалы (каталазо- и супероксидисмутазоотрицательны).
У непатогенных для человека анаэробов хемолитотрофов акцептором электронов являются неорганические кислородсодержащие соединения – нитраты, сульфаты, карбонаты.
Особую группу анаэробов составляют аэротолерантные бактерии, которые способны расти в присутствии атмосферного кислорода, но не используют его в качестве акцептора электронов (например, молочнокислые бактерии). Аэротолерантные прокариоты каталазо- и супероксиддисмутазопозитивны.
Факультативные анаэробы способны существовать как в кислородной, так и в бескислородной средах. Донорами электронов у них являются органические вещества; акцепторами электронов, в зависимости от условий среды – молекулярный или связанный в органических и неорганических соединениях кислород. Энергия факультативными анаэробами может аккумулироваться как при окислительном, так и при субстратном фосфорилировании. Как и аэробы, данная группа бактерий имеет цитохромы и ферменты антиоксидантной защиты.
Основным субстратом для получения энергии являются углеводы, которые у разных по типу дыхания хемогетеротрофных прокариот могут катаболизироваться до ацетилкоэнзима А («активированная уксусная кислота»). В качестве энергетических субстратов могут выступать липиды и белки, поскольку ацетилкоэнзим А также является одним из промежуточных продуктов их метаболизма (схема 2).
Катаболизм углеводов у хемоорганотрофных прокариот включает: (а) анаэробные процессы – гликолиз, пентозофосфатный путь и кетодезоксифосфоглюконатный путь; (б) аэробный процесс – цикл трикарбоновых кислот (цикл Кребса). Анаэробные процессы имеют место у всех прокариот, тогда как аэробный процесс характерен только для облигатных аэробов и факультативных анаэробов. В основе получения энергии анаэробными путями лежит субстратное фосфорилирование, в основе аэробного процесса – окислительное фосфорилирование.
Определение понятий.
Стерилизация, дезинфекция и антисептика являются неотъемлемыми частями современной медицинской и в особенности хирургической практики. Понимание принципов и практического применения этих методов необходимо, поскольку многие потенциально патогенные микроорганизмы способны оставаться жизнеспособными вне макроорганизма в течение длительного времени, проявлять высокую устойчивость к действию физических и химических дезинфектантов и относительно легко передаваться от одного человека к другому.
Антисептика - уничтожение или предотвращение роста патогенных или условно-патогенных микроорганизмов химическими методами. Этот термин обычно используют для обозначения наружного нанесения химического препарата на живые ткани.
Антисептик - вещество, которое угнетает рост или разрушает микроорганизм (без действия на споры бактерий). Термин является специфическим для обозначения веществ, которые используются для местного действия на живые ткани.
Асептика означает отсутствие сепсиса, но вообще этот термин используют для того, чтобы подчеркнуть отсутствие любых живых организмов. Асептические методы означают любую процедуру, предназначенную для элиминации живых организмов и предотвращения повторной контаминации ними. Современные хирургические и микробиологические методы основаны на асептических процедурах.
Биоцид - вещество, которое убивает все живые микроорганизмы, как патогенные, так и непатогенные, включая споры.
Биостат - агент, который предотвращает рост микроорганизмов, но необязательно убивает их.
Деконтаминация - удаление микроорганизмов без количественного определения. Этот термин является относительным; окончательное удаление микробов может быть осуществлено стерилизацией или дезинфекцией.
Дезинфекция - процесс, который уменьшает количество или полностью уничтожает все патогенные микроорганизмы, кроме спор.
Гермицид - вещество, которое разрушает микроорганизмы, особенно патогенные. Гермицид не разрушает споры.
Санация - метод, благодаря которому микробная контаминация уменьшается до “безопасного” уровня. Этот метод ранее использовали для “очищения” неживых объектов.
Стерилизация - использование физических факторов и (или) химических веществ для полного уничтожения или разрушения всех форм микробной жизни.
Стерилизация.
Стерилизацию определяют как разрушение или удаление (путем фильтрации) всех микроорганизмов и их спор. Стерилизацию обычно проводят с помощью тепла. Стерилизация, будучи одной из повседневных процедур в работе микробиологической лаборатории, является необходимым методом, обеспечивающим такую обработку, при которой культуры, оборудование, посуда и среды способствуют росту только необходимых микроорганизмов, тогда как другие микробы разрушаются. Различают такие виды стерилизации: прокаливание в пламени горелки, кипячение, действие текучим паром, паром под давлением в автоклаве, сухим жаром, пастеризация, тиндализация, химическая, холодная (механическая) стерилизация.
Выбор методов стерилизации.
При выборе методов стерилизации нужно учитывать следующие требования:
1. Активность: бактерицидная, спороцидная, туберкулоцидная, фунгицидная и вирусоцидная.
2. Скорость процедуры: стерилизация должна проводиться как можно более быстро.
3. Проницаемость: вещества-стерилизаторы должны проникать через упаковку и к внутренним частям инструментария.
4. Совместимость: не должны возникать изменения структуры или функции материалов, которые стерилизуют несколько раз.
5. Нетоксичность: не должно возникать угрозы для здоровья человека и состояния окружающей среды.
6. Устойчивость органического материала: эффективность стерилизации не должна снижаться в присутствия органического материала.
7. Приспособляемость: возможность использовать для больших и малых объёмов стерилизуемого материала.
8. Контроль в течение времени: цикл обработки должен легко и точно контролироваться.
9. Цена: разумная стоимость оснащения, установки и эксплуатации.
 Физические стерилизаторы
Физические стерилизаторы
Влажное тепло, которое образуется в процессе парового автоклавирования, является основным стерилизующим агентом, используемым в лабораториях клинической микробиологии. Автоклавы используют для стерилизации питательных сред, жароустойчивых материалов и обработки инфицированных отходов. Паровой стерилизатор, или автоклав, представляет собой изолированную камеру под давлением, которая использует насыщенный пар для создания высоких температур (рис. 1). Воздух удаляют из камеры замещением по массе или созданием вакуума. Наиболее часто используют автоклавы с замещением по массе. Более лёгкий пар запускают в камеру для вытеснения более тяжёлого воздуха. Кратковременная обработка паром под давлением может уничтожить бактериальные споры. Для рутинной стерилизации питательных сред и других материалов время экспозиции составляет 15 минут при 121ºС и давление - 1,5 кг на 1 квадратный сантиметр. Для инфекционных отходов время экспозиции увеличивается до 30-60 минут. Дополнительно к правильно выбранным времени и температуре, очень важным при стерилизации является прямой контакт с паром. При обработке инфекционного материала следует обеспечить максимальное проникновение пара в отходы. Такой материал необходимо обрабатывать при температуре 132ºС. Не подлежат автоклавированию антинеопластические препараты, токсичные химические вещества и радиоизотопы, которые могут не разрушиться, а также нестабильные химикаты, поскольку они под действием тепла могут испариться и распространиться по камере.
 Стерилизация сухим жаром
используется для материалов, которые невозможно стерилизовать паром в связи с возможностью повреждения или в связи с непроницаемостью материала для пара. Сухой жар менее эффективен, чем влажное тепло, и требует болеего времени экспозиции и более высоких температур. Стерилизацию сухим жаром обычно проводят в сухожаровом шкафу (рис. 2). Механизм стерилизации с помощью сухого жара является окислительным процессом. Примерами материалов, для которых используют стерилизацию сухим жаром, являются масла, порошки, острые инструменты и стеклянная посуда. Сухой жар или термическую инактивацию-стерилизацию используют как альтернативные методы обработки инфекционных отходов.
Стерилизация сухим жаром
используется для материалов, которые невозможно стерилизовать паром в связи с возможностью повреждения или в связи с непроницаемостью материала для пара. Сухой жар менее эффективен, чем влажное тепло, и требует болеего времени экспозиции и более высоких температур. Стерилизацию сухим жаром обычно проводят в сухожаровом шкафу (рис. 2). Механизм стерилизации с помощью сухого жара является окислительным процессом. Примерами материалов, для которых используют стерилизацию сухим жаром, являются масла, порошки, острые инструменты и стеклянная посуда. Сухой жар или термическую инактивацию-стерилизацию используют как альтернативные методы обработки инфекционных отходов.
Пастеризация разрушает патогенные микроорганизмы путём быстрого нагревания вещества до 71,1ºС на протяжении 15 с, что сопровождается последующим быстрым охлаждением. Пастеризация не является стерилизацией, поскольку не все микроорганизмы чувствительны к ней. Этот метод элиминировал пищевой путь передачи таких заболеваний, как туберкулез пищеварительного тракта и Q-лихорадка.
Тиндализация - это метод стерилизации прерывистым нагреванием, который может использоваться для уничтожения всех бактерий в растворах. Поскольку растущие бактерии легко гибнут при кратковременном кипячении (5 раз в течение 1 часа по 5 минут), всё, что необходимо сделать, это позволить раствору постоять на протяжении определенного времени, прежде чем тепло нарушит созревание спор с существенной потерей их устойчивости к теплу.
Фильтрация - это процесс, который используют для удаления микробов и микроскопических частей из растворов, воздуха и других газов. Наиболее часто стерилизацию путем фильтрации в лаборатории используют для обработки диагностических препаратов, питательных сред, тканевых культуральных сред, сывороток, растворов, которые содержат компоненты сыворотки. Другим общепринятым применением фильтрации является стерилизация воздуха и газов. Пластиковые или бумажные мембранные фильтры, которые различают по диаметру пор (примерно от 12 до 0,22 μм) и используют для механического разделения, служат и для сбора микробов из жидкостей для микроскопического изучения или культивирования прямо на фильтре, когда его помещают на поверхность, пропитанную питательной средой.
Ультрафиолетовое облучение является видом электромагнитной волновой радиации, которая действует на клеточную нуклеиновую кислоту. Микроорганизмы высокочувствительны к действию ультрафиолетовых лучей с длиной волны 254 нм. Ультрафиолет наиболее широко используют для уничтожения микроорганизмов, находящихся в воздухе или на каких-либо поверхностях. Другим применением является холодная стерилизация определенных химикатов и пластика для фармацевтических целей, стерилизация сыворотки для клеточных культур и дезинфекция воды. Существенным недостатком ультрафиолетового облучения в качестве стерилизатора является его неспособность к проникновению внутрь материалов.
Ионизирующее излучение в электромагнитном спектре летально действует на микроорганизмы. Этот спектр включает микроволны, γ-лучи, рентгеновские лучи и поток электронов. Летальный эффект от ионизирующего излучения возникает вследствие прямого действия на молекулу-мишень, в результате чего энергия переносится в молекулу; и вследствие косвенного действия - диффузии радикалов.
Ультразвуковая энергия с низкой частотой инактивирует микроорганизмы в водных растворах. Физический эффект обработки ультразвуком возникает вследствие кавитации. Ультразвуковые очистители и другие приборы часто используют для очистки инструментов, но не считают стерилизаторами. Однако комбинирование ультразвука с химической обработкой убивает микроорганизмы.
Химические стерилизаторы
2 % глютаровый альдегид в качестве жидкого химического стерилизатора ранее широко применяли для обработки медицинского и хирургического материала, который невозможно стерилизовать нагреванием или облучением. Глютаровый альдегид также используют при приготовлении вакцин.
Дезинфекция.
Дезинфекцию можно проводить химическими методами или кипячением. Кипячение является эффективным методом дезинфекции инструментария, например, игл и шприцев, если нет автоклава. Предварительно очищенный медицинский инструментарий следует кипятить 20 минут. Химическую дезинфекцию используют для чувствительного к действию тепла оборудования, которое может повредить высокая температура. Широко используют такие химические дезинфектанты, как компонента хлора, этиловый и изопропиловый спирт, четвертичные компоненты аммония и глютаровый альдегид.
Химические дезинфектанты.
Спирт (этиловый и изопропиловый) , растворённый в воде до концентрации 60-85 %, очень эффективен при дезинфекции. Спирты имеют бактерицидное, фунгицидное и туберкулоцидное действие, но не влияют на споры. Этиловый спирт имеет более широкий спектр вирусоцидной активности, чем изопропиловый, поэтому он более эффективно действует на липофильные и гидрофильные вирусы.
Раствор 37 % формальдеида , который называют формалином, можно использовать в качестве стерилизатора, тогда как его концентрации 3-8 % можно использовать в качестве дезинфектантов.
Фенол в чистом виде не используют в качестве дезинфектанта в связи с его токсичностью, способностью индуцировать развитие опухолей и коррозии. Дериваты фенола, в которых функциональная группа (хлор, бром, алкил, бензил, фенил, амил) замещает один из атомов водорода в ароматическом кольце, широко используют в качестве дезинфектантов. Подобное замещение уменьшает недостатки фенола. Компоненты фенола убивают микробы благодаря инактивации ферментных систем, преципитации белков и нарушению клеточной стенки и мембраны. Обычно используют концентрации 2-5 %, более низкая концентрация требует более длительной экспозиции.
Галогены. Только хлор и йод используют для дезинфекции в лабораторной практике. В связи с тем, что хлор является мощным окислителем, считают, что он убивает микробы путем окисления. Считают, что йод убивает микроорганизмы путём реакции с N-H и S-H группами аминокислот, а также с фенольной группой аминокислоты тирозина и углерод-углеродными двойными связями ненасыщенных жирных кислот. Обычная обработка включает распыление 2-5 % раствора формальдегида в присутствии пара при температуре 60-80ºС.
Антисептика.
Антисептики можно обнаружить в микробиологических лабораториях, прежде всего, в веществах, которые используют для мытья рук. В тех случаях, когда медицинский персонал оказывает неотложную помощь пациентам с использованием веществ, содержащих антибактериальные агенты, это уменьшает количество госпитальных инфекций. Наиболее распространёнными химическими соединениями, содержащимися в веществах для мытья рук, являются спирты, хлоргексидина глюконат, йодофоры, хлороксайленол и триклозан.
Традиционными методами обработки отходов и мусора являются сжигание и стерилизация паром.
Сжигание является методом выбора для обработки отходов и мусора. Этот метод делает отходы неинфекционными, а также изменяет их форму и размеры. Стерилизация является эффективным методом обработки отходов, но она не изменяет их формы. Стерилизация паром в автоклаве при 121ºС в течение минимум 15 минут уничтожает все формы микробной жизни, включая большое количество бактериальных спор. Этот тип полной стерилизации также можно провести с использованием сухого жара при температуре 160-170ºС на протяжении 2-4 часов. Однако следует убедиться, что сухой жар контактирует со стерилизуемым материалом. Поэтому бутылки, которые содержат жидкость, должны быть неплотно закрыты пробками или ватными тампонами для того, чтобы пар и жар могли обмениваться с воздухом в бутылках. Биологически опасные контейнеры, содержащие отходы, следует плотно завязать. Простерилизованный биологически опасный материал нужно запечатать в соответствующие контейнеры с этикетками.
Стерилизация паром (в автоклаве). Инфекционный мусор считают деконтаминированным при уменьшении в 6 lg раз количества вегетативных бактерий, грибов, микобактерий и вирусов, содержащих липиды, и в 4 lg раза - бактериальных эндоспор.
Питание бактерий.
Питание. Под питанием бактериальной клетки следует понимать процесс поглощения и усвоения пластического материала и энергии в результате преобразовательных реакций . Типы питания прокариот сложны и разнообразны. Они различаются в зависимости от способа поступления питательных веществ внутрь бактериальной клетки, источников углерода и азота, способа получения энергии, природы доноров электронов.
Транспорт питательных веществ внутрь клетки может осуществляться 3 механизмами: пассивной диффузией, облегчённой диффузией и активным транспортом.
Пассивная диффузия является неспецифическим энергозависимым процессом, осуществляемым по градиенту концентрации веществ (вещество из среды с большей своей концентрацией пассивно, согласно законам осмоса, поступает в среду с меньшей концентрацией). Пассивной диффузией внутрь бактериальной клетки поступает ограниченное количество веществ, некоторые ионы, моносахара. Скорость переноса веществ при пассивной диффузии незначительна и зависит от липофильности и размеров транспортирующихся молекул.
Облегчённая диффузия представляет собой энергонезависимый транспорт веществ по градиенту концентрации при помощи ферментов пермеаз. Пермеазы – это специфические мембранные белки, способствующие прохождению веществ через цитоплазматическую мембрану. Пермеаза фиксирует на себе молекулу переносимого вещества, вместе с которым пеодолевает цитоплазматическую мембрану, после чего комплекс «вещество – пермеаза» диссоциирует. Освободившаяся пермеаза используется для проведения других молекул. У прокариотов облегчённой диффузией внутрь клетки поступает только глицерин. При этом внутриклеточная концентрация глицерина соответствует таковой вне клетки. Облегчённая диффузия наиболее характерна для микроорганизмов-эукариот.
Активный транспорт – это энергозависимый перенос веществ внутрь клетки против градиента концентрации при помощи специфических ферментов. Активным транспортом в бактериальную клетку поступает подавляющее большинство веществ (ионы, углеводы, аминокислоты, липиды и др.). Активный транспорт может осуществляться: (1) без химической модификации переносимого вещества; (2) с химической модификацией.
Бактерии являются прокариотическими микроорганизмами клеточного строения. Их размеры - от 0,1 до 30 мкм. Микробы чрезвычайно распространены. Они живут в почве, воздухе, воде, в снегах и даже горячих источниках, на теле животных, а также внутри живых организмов, в том числе в организме человека.
Распределение бактерий на виды идет с учетом нескольких критериев, среди которых чаще всего принимают во внимание форму микроорганизмов и их пространственное размещение. Так, по форме клеток бактерии делятся на:
Коки - микро-, дипло-, стрепто-, стафилококки, а также сарцины;
Палочковидные - монобактерии, диплобактерии и стрептобактерии;
Извитые виды - вибрионы и спирохеты.
Определитель Берджи систематизирует все известные бактерии по нашедшим в практической бактериологии наибольшее распространение принципам идентификации бактерий, основанным на различиях в строении клеточной стенки и отношении к окраске по Граму. Описание бактерий даётся по группам (секциям), в состав которых включены семейства, роды и виды; в некоторых случаях в состав групп входят классы и порядки. Патогенные для человека бактерии входят в небольшое число групп.
Определитель выделяет четыре основных категории бактерий -
Gracillicutes [от лат. gracilis, изящный, тонкий, + cutis, кожа] - виды с тонкой клеточной стенкой, окрашивающиеся грамотрицательно ;
firmicutes [от лат. flrmus, крепкий, + cutis, кожа] - бактерии с толстой клеточной стенкой, окрашивающиеся грамположительно ;
Tenericutes [от лат. tener, нежный, + cutis, кожа] - бактерии, лишённые клеточной стенки (микоплазмы и прочие представители класса Mollicutes)
Mendosicutes [от лат. mendosus, неправильный, + cutis, кожа] - архебактерии (метан- и сульфатредуцирующие, галофильные, термофильные и архебактерии, лишённые клеточной стенки).
Группа 2 определителя Берджи. Аэробные и микроаэрофильные подвижные извитые и изогнутые грамотрицательные бактерии. Патогенные для человека виды входят в роды Campylobacter, Helicobacters Spirillum.
Группа 3 определителя Берджи. Неподвижные (редко подвижные) грамотрицательные бактерии. Не содержит патогенные виды.
Группа 4 определителя Берджи. Грамотрицательные аэробные и микроаэрофильные палочки и кокки. Патогенные для человека виды включены в состав семейств Legionellaceae, Neisseriaceae и Pseudomonada-сеае, в группу входят также патогенные и условно-патогенные бактерии родов Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Francisella, Kingella и Moraxella.
Группа 5 определителя Берджи. Факультативно анаэробные грамотрицательные палочки. Группа образована тремя семействами - Enterobacteriaceae, Vibrionaceae и Pasteurellaceae, каждое из которых включает патогенные виды, а также патогенные и условно-патогенные бактерии родов Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella и Streptobacillus.
Группа 6 определителя Берджи. Грамотрицательные анаэробные прямые, изогнутые и спиральные бактерии. Патогенные и условно-патогенные виды входят в состав родов Bacteroides, Fusobacterium, Porphoromonas и Prevotelta.
Группа 7 определителя Берджи. Бактерии, осуществляющие диссимиляционное восстановление сульфата или серы Не включает патогенные виды.
Группа 8 определителя Берджи. Анаэробные грамотрицательные кокки. Включает условно-патогенные бактерии poда Veillonella.
Группа 9 определителя Берджи. Риккетсии и хламидии. Три семейства - Rickettsiaceae, Bartonellaceae и Chlamydiaсеае, каждое из которых содержит патогенные для человека виды.
Группы 10 и 11 определителя Берджи включают анокси- и оксигенные фототрофные бактерии, не патогенные для человека.
Группа 12 определителя Берджи. Аэробные хемолитотрофные бактерии и родственные организмы. Объединяет серо- железо- и марганецокисляющие и нитрифицирующие бактерии, не вызывающие поражения у человека.
Группы 13 и 14 определителя Берджи включают почкующиеся и/или обладающие выростами бактерии и бактерии образующие футляры. Представлены свободноживущими видами, не патогенными для человек;
Группы 15 и 16 определителя Берджи объединяют скользящие бактерии, не образующие плодовые тела и образующие их. Группы не включают виды, патогенные для человека.
Группа 17 определителя Берджи. Грамположительные кокки. Включает условно-патогенные виды родов Enterococcus Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus.
Группа 18 определителя Берджи. Спорообразующие грамположительные палочки и кокки. Включает патогенные, условно-патогенные палочки родов Clostridium и Bacillus.
Группа 19 определителя Берджи. Споронеобразующие грамположительные палочки правильной формы. Включая условно-патогенные виды родов Erysipelothrix и Listeria.
Группа 20 определителя Берджи. Споронеобразующие грамположительные палочки неправильной формы. В состав группы входят патогенные и условно-патогенные виды родов Actinomyces, Corynebacterium Gardnerella, Mobiluncus и др.
Группа 21 определителя Берджи. Микобактерии. Включает единственный род Mycobacterium, объединяющий патогенные и условно-патогенные виды.
Группы 22-29. Актиномицеты. Среди многочисленных видов лишь нокардиоформные актиномицеты (группа 22) родов Gordona, Nocardia, Rhodococcus, Tsukamurella, Jonesia, Oerskovi и Terrabacter способны вызывать поражения у человека.
Группа 30 определителя Берджи. Микоплазмы. Патогенны для человека виды, включённые в состав рода Acholeplasma, Mycoplasma и Ureaplasma.
Остальные группы определителя Берджи - метаногенные бактерии (31), сульфатредуцируюшие бактерии (32 экстремально галофильные аэробные архебактерии (33), архебактерии, лишённые клеточно стенки (34), экстремальные термофилы и гипертермофилы, метаболизируюшие серу (35) - не содержат патогенные для человека виды.
Бактерии — самая древняя группа организмов из ныне существующих на Земле. Первые бактерии появились, вероятно, более 3,5 млрд лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. Поскольку это были первые представители живой природы, их тело имело примитивное строение.
Со временем их строение усложнилось, но и поныне бактерии считаются наиболее примитивными одноклеточными организмами. Интересно, что некоторые бактерии и сейчас ещё сохранили примитивные черты своих древних предков. Это наблюдается у бактерий, обитающих в горячих серных источниках и бескислородных илах на дне водоёмов.
Большинство бактерий бесцветно. Только немногие окрашены в пурпурный или в зелёный цвет. Но колонии многих бактерий имеют яркую окраску, которая обусловливается выделением окрашенного вещества в окружающую среду или пигментированием клеток.
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель 17 века, впервые создавший совершенную лупу-микроскоп, увеличивающую предметы в 160-270 раз.
Бактерии относят к прокариотам и выделяют в отдельное царство — Бактерии.
Форма тела
Бактерии — многочисленные и разнообразные организмы. Они различаются по форме.
| Название бактерии | Форма бактерии | Изображение бактерии |
| Кокки | Шарообразная | |
| Бацилла |  | Палочковидная |
| Вибрион | Изогнутая в виде запятой | |
| Спирилла |  | Спиралевидная |
| Стрептококки |  | Цепочка из кокков |
| Стафилококки |  | Грозди кокков |
| Диплококки | Две круглые бактерии, заключённые в одной слизистой капсуле |
Способы передвижения
Среди бактерий есть подвижные и неподвижные формы. Подвижные передвигаются за счёт волнообразных сокращений или при помощи жгутиков (скрученные винтообразные нити), которые состоят из особого белка флагеллина. Жгутиков может быть один или несколько. Располагаются они у одних бактерий на одном конце клетки, у других — на двух или по всей поверхности.

Но движение присуще и многим иным бактериям, у которых жгутики отсутствуют. Так, бактерии, покрытые снаружи слизью, способны к скользящему движению.
У некоторых лишённых жгутиков водных и почвенных бактерий в цитоплазме имеются газовые вакуоли. В клетке может быть 40-60 вакуолей. Каждая из них заполнена газом (предположительно — азотом). Регулируя количество газа в вакуолях, водные бактерии могут погружаться в толщу воды или подниматься на её поверхность, а почвенные бактерии — передвигаться в капиллярах почвы.
Место обитания
В силу простоты организации и неприхотливости бактерии широко распространены в природе. Бактерии обнаружены везде: в капле даже самой чистой родниковой воды, в крупинках почвы, в воздухе, на скалах, в полярных снегах, песках пустынь, на дне океана, в добытой с огромной глубины нефти и даже в воде горячих источников с температурой около 80ºС. Обитают они на растениях, плодах, у различных животных и у человека в кишечнике, ротовой полости, на конечностях, на поверхности тела.
Бактерии — самые мелкие и самые многочисленные живые существа. Благодаря малым размерам они легко проникают в любые трещины, щели, поры. Очень выносливы и приспособлены к различным условиям существования. Переносят высушивание, сильные холода, нагревание до 90ºС, не теряя при этом жизнеспособность.
Практически нет места на Земле, где не встречались бы бактерии, но в разных количествах. Условия жизни бактерий разнообразны. Одним из них необходим кислород воздуха, другие в нём не нуждаются и способны жить в бескислородной среде.
В воздухе: бактерии поднимаются в верхние слои атмосферы до 30 км. и больше.
Особенно много их в почве. В 1 г. почвы могут содержаться сотни миллионов бактерий.
В воде: в поверхностных слоях воды открытых водоёмов. Полезные водные бактерии минерализуют органические остатки.
В живых организмах: болезнетворные бактерии попадают в организм из внешней среды, но лишь в благоприятных условиях вызываю заболевания. Симбиотические живут в органах пищеварения, помогая расщеплять и усваивать пищу, синтезируют витамины.
Внешнее строение
Клетка бактерии одета особой плотной оболочкой — клеточной стенкой, которая выполняет защитную и опорную функции, а также придаёт бактерии постоянную, характерную для неё форму. Клеточная стенка бактерии напоминает оболочку растительной клетки. Она проницаема: через неё питательные вещества свободно проходят в клетку, а продукты обмена веществ выходят в окружающую среду. Часто поверх клеточной стенки у бактерий вырабатывается дополнительный защитный слой слизи — капсула. Толщина капсулы может во много раз превышать диаметр самой клетки, но может быть и очень небольшой. Капсула — не обязательная часть клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она предохраняет бактерию от высыхания.
На поверхности некоторых бактерий имеются длинные жгутики (один, два или много) или короткие тонкие ворсинки. Длина жгутиков может во много раз превышать разметы тела бактерии. С помощью жгутиков и ворсинок бактерии передвигаются.

Внутреннее строение
Внутри клетки бактерии находится густая неподвижная цитоплазма. Она имеет слоистое строение, вакуолей нет, поэтому различные белки (ферменты) и запасные питательные вещества размещаются в самом веществе цитоплазмы. Клетки бактерий не имеют ядра. В центральной части их клетки сконцентрировано вещество, несущее наследственную информации. Бактерии, — нуклеиновая кислота — ДНК. Но это вещество не оформлено в ядро.
Внутренняя организация бактериальной клетки сложна и имеет свои специфические особенности. Цитоплазма отделяется от клеточной стенки цитоплазматической мембраной. В цитоплазме различают основное вещество, или матрикс, рибосомы и небольшое количество мембранных структур, выполняющих самые различные функции (аналоги митохондрий, эндоплазматической сети, аппарата Гольджи). В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Гранулы могут состоять из соединений, которые служат источником энергии и углерода. В бактериальной клетке встречаются и капельки жира.
В центральной части клетки локализовано ядерное вещество — ДНК, не отграниченная от цитоплазмы мембраной. Это аналог ядра — нуклеоид. Нуклеоид не обладает мембраной, ядрышком и набором хромосом.
Способы питания
У бактерий наблюдаются разные способы питания. Среди них есть автотрофы и гетеротрофы. Автотрофы — организмы, способные самостоятельно образовывать органические вещества для своего питания.
Растения нуждаются в азоте, но сами усваивают азот воздуха не могут. Некоторые бактерии соединяют содержащиеся в воздухе молекулы азота с другими молекулами, в результате чего получаются вещества, доступные для растений.
Эти бактерии поселяются в клетках молодых корней, что приводит к образованию на корнях утолщений, называемых клубеньками. Такие клубеньки образуются на корнях растений семейства бобовых и некоторых других растений.

Корни дают бактериям углеводы, а бактерии корням — такие содержащие азот вещества, которые могут быть усвоены растением. Их сожительство взаимовыгодно.
Корни растений выделяют много органических веществ (сахара, аминокислоты и другие), которыми питаются бактерии. Поэтому в слое почвы, окружающем корни, поселяется особенно много бактерий. Эти бактерии превращают отмершие остатки растений в доступные для растения вещества. Этот слой почвы называют ризосферой.
Существует несколько гипотез о проникновении клубеньковых бактерий в ткани корня:
- через повреждения эпидермальной и коровой ткани;
- через корневые волоски;
- только через молодую клеточную оболочку;
- благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты;
- благодаря стимуляции синтеза В-индолилуксусной кислоты из триптофана, всегда имеющегося в корневых выделениях растений.
Процесс внедрения клубеньковых бактерий в ткань корня состоит из двух фаз:
- инфицирование корневых волосков;
- процесс образования клубеньков.
В большинстве случаев внедрившаяся клетка, активно размножается, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина.

Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. В период функционирования клубеньки обычно плотные. К моменту проявления оптимальной активности клубеньки приобретают розовую окраску (благодаря пигменту легоглобину). Фиксировать азот способны лишь те бактерии, которые содержат легоглобин.
Бактерии клубеньков создают десятки и сотни килограммов азотных удобрений на гектаре почвы.
Обмен веществ
Бактерии отличаются друг от друга обменом веществ. У одних он идёт при участии кислорода, у других — без его участия.
Большинство бактерий питается готовыми органическими веществами. Лишь некоторые из них (сине-зелёные, или цианобактерии), способны создавать органические вещества из неорганических. Они сыграли важную роль в накоплении кислорода в атмосфере Земли.
Бактерии впитывают вещества извне, разрывают их молекулы на части, из этих частей собирают свою оболочку и пополняют своё содержимое (так они растут), а ненужные молекулы выбрасывают наружу. Оболочка и мембрана бактерии позволяет ей впитывать только нужные вещества.

Если бы оболочка и мембрана бактерии были полностью непроницаемыми, в клетку не попали бы никакие вещества. Если бы они были проницаемыми для всех веществ, содержимое клетки перемешалось бы со средой — раствором, в которой обитает бактерия. Для выживания бактерии необходима оболочка, которая нужные вещества пропускает, а ненужные — нет.
Бактерия поглощает находящиеся близ неё питательные вещества. Что происходит потом? Если она может самостоятельно передвигаться (двигая жгутик или выталкивая назад слизь), то она перемещается, пока не найдёт необходимые вещества.

Если она двигаться не может, то ждёт, пока диффузия (способность молекул одного вещества проникать в гущу молекул другого вещества) не принесёт к ней необходимые молекулы.
Бактерии в совокупности с другими группами микроорганизмов выполняют огромную химическую работу. Превращая различные соединения, они получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий разнообразны.
Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют за счёт неорганических соединений. Они называются автотрофами. Автотрофные бактерии способны синтезировать органические вещества из неорганических. Среди них различают:
Хемосинтез
Использование лучистой энергии — важнейший, но не единственный путь создания органического вещества из углекислого газа и воды. Известны бактерии, которые в качестве источника энергии для такого синтеза используют не солнечный свет, а энергию химических связей, происходящих в клетках организмов при окислении некоторых неорганических соединений — сероводорода, серы, аммиака, водорода, азотной кислоты, закисных соединений железа и марганца. Образованное с использованием этой химической энергии органическое вещество они используют для построения клеток своего тела. Поэтому такой процесс называют хемосинтезом.
Важнейшую группу хемосинтезирующих микроорганизмов составляют нитрифицирующие бактерии. Эти бактерии живут в почве и осуществляют окисление аммиака, образовавшегося при гниении органических остатков, до азотной кислоты. Последняя, реагирует с минеральными соединениями почвы, превращаются в соли азотной кислоты. Этот процесс проходит в две фазы.
Железобактерии превращают закисное железо в окисное. Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Некоторые микроорганизмы существуют за счёт окисления молекулярного водорода, обеспечивая тем самым автотрофный способ питания.
Характерной особенностью водородных бактерий является способность переключаться на гетеротрофный образ жизни при обеспечении их органическими соединениями и отсутствии водорода.
Таким образом, хемоавтотрофы являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения, а не берут их в готовом виде от других организмов, как гетеротрофы. От фототрофных растений хемоавтотрофные бактерии отличаются полной независимостью от света как источника энергии.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты — бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка — карбоновые кислоты), а у зелёных растений — вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Спорообразование
Внутри бактериальной клетки образуются споры. В процессе спорообразования бактериальная клетка претерпевает ряд биохимических процессов. В ней уменьшается количество свободной воды, снижается ферментативная активность. Это обеспечивает устойчивость спор к неблагоприятным условиям внешней среды (высокой температуре, высокой концентрации солей, высушиванию и др.). Спорообразование свойственно только небольшой группе бактерий.

Споры — не обязательная стадия жизненного цикла бактерий. Спорообразование начинается лишь при недостатке питательных веществ или накоплении продуктов обмена. Бактерии в виде спор могут длительное время находиться в состоянии покоя. Споры бактерий выдерживают продолжительное кипячение и очень длительное проммораживание. При наступлении благоприятных условий спора прорастает и становится жизнеспособной. Спора бактерий — это приспособление к выживанию в неблагоприятных условиях.
Размножение
Размножаются бактерии делением одной клетки на две. Достигнув определённого размера, бактерия делится на две одинаковые бактерии. Затем каждая из них начинает питаться, растёт, делится и так далее.

После удлинения клетки постепенно образуется поперечная перегородка, а затем дочерние клетки расходятся; у многих бактерий в определённых условиях клетки после деления остаются связанными в характерные группы. При этом в зависимости от направления плоскости деления и числа делений возникают разные формы. Размножение почкованием встречается у бактерий как исключение.
При благоприятных условиях деление клеток у многих бактерий происходит через каждые 20-30 минут. При таком быстром размножении потомство одной бактерии за 5 суток способно образовать массу, которой можно заполнить все моря и океаны. Простой подсчёт показывает, что за сутки может образоваться 72 поколения (720 000 000 000 000 000 000 клеток). Если перевести в вес — 4720 тонн. Однако в природе этого не происходит, так как большинство бактерий быстро погибают под действием солнечного света, при высушивании, недостатке пищи, нагревании до 65-100ºС, в результате борьбы между видами и т.д.
Бактерия (1), поглотившая достаточно пищи, увеличивается в размерах (2) и начинает готовиться к размножению (делению клетки). Её ДНК (у бактерии молекула ДНК замкнута в кольцо) удваивается (бактерия производит копию этой молекулы). Обе молекулы ДНК (3,4) оказываются, прикреплены к стенке бактерии и при удлинении бактерии расходятся в стороны (5,6). Сначала делится нуклеотид, затем цитоплазма.
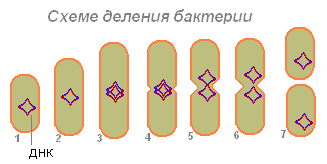
После расхождения двух молекул ДНК на бактерии появляется перетяжка, которая постепенно разделяет тело бактерии на две части, в каждой из которых есть молекула ДНК (7).
Бывает (у сенной палочки), две бактерии слипаются, и между ними образуется перемычка (1,2).

По перемычке ДНК из одной бактерии переправляется в другую (3). Оказавшись в одной бактерии, молекулы ДНК сплетаются, слипаются в некоторых местах (4), после чего обмениваются участками (5).
Роль бактерий в природе
Круговорот
Бактерии — важнейшее звено общего круговорота веществ в природе. Растения создают сложные органические вещества из углекислого газа, воды и минеральных солей почвы. Эти вещества возвращаются в почву с отмершими грибами, растениями и трупами животных. Бактерии разлагают сложные вещества на простые, которые снова используют растения.
Бактерии разрушают сложные органические вещества отмерших растений и трупов животных, выделения живых организмов и разные отбросы. Питаясь этими органическими веществами, сапрофитные бактерии гниения превращают их в перегной. Это своеобразные санитары нашей планеты. Таким образом, бактерии активно участвуют в круговороте веществ в природе.
Почвообразование
Поскольку бактерии распространены практически повсеместно и встречаются в огромном количестве, они во многом определяют различные процессы, происходящие в природе. Осенью опадают листья деревьев и кустарников, отмирают надземные побеги трав, опадают старые ветки, время от времени падают стволы старых деревьев. Всё это постепенно превращается в перегной. В 1 см 3 . поверхностного слоя лесной почвы содержатся сотни миллионов сапрофитных почвенных бактерий нескольких видов. Эти бактерии превращают перегной в различные минеральные вещества, которые могут быть поглощены из почвы корнями растений.
Некоторые почвенные бактерии способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Эти азотофиксирующие бактерии живут самостоятельно или поселяются в корнях бобовых растений. Проникнув в корни бобовых, эти бактерии вызывают разрастание клеток корней и образование на них клубеньков.
Эти бактерии выделяют азотные соединения, которые используют растения. От растений бактерии получают углеводы и минеральные соли. Таким образом, между бобовым растением и клубеньковыми бактериями существует тесная связь, полезная как одному, так и другому организму. Это явление носит название симбиоза.
Благодаря симбиозу с клубеньковыми бактериями бобовые растения обогащают почву азотом, способствуя повышению урожая.
Распространение в природе
Микроорганизмы распространены повсеместно. Исключение составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни низкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни жёсткое облучение атомных реакторов не мешают существованию и развитию микрофлоры. Все живые существа постоянно взаимодействуют с микроорганизмами, являясь часто не только их хранилищами, но и распространителями. Микроорганизмы — аборигены нашей планеты, активно осваивающие самые невероятные природные субстраты.
Микрофлора почвы
Количество бактерий в почве чрезвычайно велико — сотни миллионов и миллиардов особей в 1 грамме. В почве их значительно больше, чем в воде и воздухе. Общее количество бактерий в почвах меняется. Количество бактерий зависит от типа почв, их состояния, глубины расположения слоёв.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20-100 клеток в каждой). Часто они развиваются в толщах сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Микрофлора почвы очень разнообразна. Здесь встречаются разные физиологические группы бактерий: бактерии гниения, нитрифицирующие, азотфиксирующие, серобактерии и др. среди них есть аэробы и анаэробы, споровые и не споровые формы. Микрофлора — один из факторов образования почв.
Областью развития микроорганизмов в почве является зона, примыкающая к корням живых растений. Её называют ризосферой, а совокупность микроорганизмов, содержащихся в ней, — ризосферной микрофлорой.
Микрофлора водоёмов
Вода — природная среда, где в большом количестве развиваются микроорганизмы. Основная масса их попадает в воду из почвы. Фактор, определяющий количество бактерий в воде, наличие в ней питательных веществ. Наиболее чистыми являются воды артезианских скважин и родниковые. Очень богаты бактериями открытые водоёмы, реки. Наибольшее количество бактерий находится в поверхностных слоях воды, ближе к берегу. При удалении от берега и увеличении глубины количество бактерий уменьшается.
Чистая вода содержит 100-200 бактерий в 1 мл., а загрязнённая — 100-300 тыс. и более. Много бактерий в донном иле, особенно в поверхностном слое, где бактерии образуют плёнку. В этой плёнке много серо- и железобактерий, которые окисляют сероводород до серной кислоты и тем самым предотвращают замор рыбы. В иле больше спороносных форм, в то время как в воде преобладают неспороносные.
По видовому составу микрофлора воды сходна с микрофлорой почвы, но встречаются и специфические формы. Разрушая различные отбросы, попавшие в воду, микроорганизмы постепенно осуществляют так называемое биологическое очищение воды.
Микрофлора воздуха
Микрофлора воздуха менее многочисленна, чем микрофлора почвы и воды. Бактерии поднимаются в воздух с пылью, некоторое время могут находиться там, а затем оседают на поверхность земли и гибнут от недостатка питания или под действием ультрафиолетовых лучей. Количество микроорганизмов в воздухе зависит от географической зоны, местности, времени года, загрязнённостью пылью и др. каждая пылинка является носителем микроорганизмов. Больше всего бактерий в воздухе над промышленными предприятиями. Воздух сельской местности чище. Наиболее чистый воздух над лесами, горами, снежными пространствами. Верхние слои воздуха содержат меньше микробов. В микрофлоре воздуха много пигментированных и спороносных бактерий, которые более устойчивы, чем другие, к ультрафиолетовым лучам.
Микрофлора организма человека
Тело человека, даже полностью здорового, всегда является носителем микрофлоры. При соприкосновении тела человека с воздухом и почвой на одежде и коже оседают разнообразные микроорганизмы, в том числе и патогенные (палочки столбняка, газовой гангрены и др.). Наиболее часто загрязняются открытые части человеческого тела. На руках обнаруживают кишечные палочки, стафилококки. В ротовой полости насчитывают свыше 100 видов микробов. Рот с его температурой, влажностью, питательными остатками — прекрасная среда для развития микроорганизмов.
Желудок имеет кислую реакцию, поэтому основная масса микроорганизмов в нём гибнет. Начиная с тонкого кишечника реакция становится щелочной, т.е. благоприятной для микробов. В толстых кишках микрофлора очень разнообразна. Каждый взрослый человек выделяет ежедневно с экскрементами около 18 млрд. бактерий, т.е. больше особей, чем людей на земном шаре.
Внутренние органы, не соединяющиеся с внешней средой (мозг, сердце, печень, мочевой пузырь и др.), обычно свободны от микробов. В эти органы микробы попадают только во время болезни.
Бактерии в круговороте веществ
Микроорганизмы вообще и бактерии в частности играют большую роль в биологически важных круговоротах веществ на Земле, осуществляя химические превращения, совершенно недоступные ни растениям, ни животным. Различные этапы круговорота элементов осуществляются организмами разного типа. Существование каждой отдельной группы организмов зависит от химического превращения элементов, осуществляемого другими группами.
Круговорот азота
Циклическое превращение азотистых соединений играет первостепенную роль в снабжении необходимыми формами азота различных по пищевым потребностям организмов биосферы. Свыше 90% общей фиксации азота обусловлено метаболической активностью определённых бактерий.
Круговорот углерода
Биологическое превращение органического углерода в углекислый газ, сопровождающееся восстановлением молекулярного кислорода, требует совместной метаболической активности разнообразных микроорганизмов. Многие аэробные бактерии осуществляют полное окисление органических веществ. В аэробных условиях органические соединения первоначально расщепляются путём сбраживания, а органические конечные продукты брожения окисляются далее в результате анаэробного дыхания, если имеются неорганические акцепторы водорода (нитрат, сульфат или СО 2).
Круговорот серы
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
Круговорот железа
В некоторых водоёмах с пресной водой содержатся в высоких концентрациях восстановленные соли железа. В таких местах развивается специфическая бактериальная микрофлора — железобактерии, окисляющие восстановленное железо. Они участвуют в образовании болотных железных руд и водных источников, богатых солями железа.
Бактерии являются самыми древними организмами, появившимися около 3,5 млрд. лет назад в архее. Около 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы.
Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они - первые организмы, появившиеся на Земле.
Бактерии - это одноклеточные, бесхлорофильные организмы, вегетативно размножающиеся делением, реже перешнурованием, иногда образующие внутриклеточные споры.
Величина бактерий измеряется в микронах и за редкими исключениями лежит в пределах от 0,06-0,3 до 3-5 μ. В капле воды могут свободно поместиться несколько сот миллионов бактерий.
Форма бактериальной клетки довольно однообразна. Известны три основные формы бактерий: круглая, палочковидная и извитая с многочисленными и незаметными переходами между ними. Антон Де-Бари образно сравнивал их с бильярдным шаром, карандашом и пробочником.
Кокками называются бактерии, имеющие круглую форму. Они различаются размерами и взаимным расположением. Кокки, соединенные попарно, носят название диплококков, соединенные же в виде ожерелья называются стрептококками. При делении, чередующемся в двух взаимно-перпендикулярных направлениях, образуются тетракокки. Если деление правильно и повторяется в трех взаимно-перпендикулярных направлениях, то образуются соединения клеток в виде пакетов - это так называемые сардины. Делясь в разных направлениях без особой правильности, кокки образуют беспорядочные скопления, напоминающие кисть винограда. Они называются стафилококками.
Бактерии палочковидной формы несколько более разнообразны по своему внешнему виду. Они могут быть с усеченными или закругленными концами, цилиндрическими, бочковидными или лимоновидными и как бы с перетяжкой посередине, часто эллипсоидальными, отличаясь лишь своими размерами в ширину и длину. Палочки могут быть прямыми, изогнутыми, одиночными, сцепленными попарно или цепочкой, короткими или сильно вытянутыми. Палочковидные бактерии, у которых длина в два раза и более превосходит ширину, называются бациллами; если же разница между длиной и шириной невелика, то их называют бактериями.
Бактерии извитой формы различаются не только по длине и толщине, но и по количеству и характеру завитков. Слегка изогнутые бактерии (завиток не превышает 1/4 оборота спирали) называются вибрионами, бактерии, имеющие один или несколько больших правильных завитков, - спириллами. Длинные и тонкие бактерии извитой формы с многочисленными мелкими завитками, иногда с крупными искривлениями всей нити, называются спирохетами.
Строение бактерий
По простоте своей организации и ничтожным размерам бактерии принадлежат к наиболее элементарным существам и стоят на самых низких ступенях жизни. Несмотря на огромные успехи науки и техники, еще не все вопросы строения бактерий разрешены.
Тело бактерий состоит из оболочки и протоплазм этического содержимого, пропитанного клеточным соком. Оболочка бактерий тонка, бесцветна, структура ее не различима в микроскоп. Для того чтобы видеть ее, прибегают к искусственным методам обработки. Оболочка лежит в основе внешней формы клетки и, по-видимому, является известной защитой в неблагоприятных условиях. Свободно облекая содержимое клетки, она благодари своей упругости допускает свободное движение бактерий, нередко сопровождающееся оживленными движениями всего тела.
Наружные слои оболочки, поглощая воду, нередко разбухают и образуют студенистую клейкую массу, достигающую заметных размеров. По мере ослизнения наружных слоев оболочка беспрерывно возобновляется за счет протоплазмы. Остудневшая клейкая оболочка носит название капсулы. Интенсивность образования слизи и капсул зависит от особенностей питания и иногда может быть весьма значительной. Слизистая капсула образуется не только около каждой клетки в отдельности, но и у многих клеток, связанных в одну колонию и заключенных как бы в общую капсулу. Такие слизистые колонии бактерий называются зооглеями . Образование капсул свойственно не всем видам бактерий.
Движение бактерий
Способность к самопроизвольному движению присуща лишь некоторым группам бактерий. Передвигаются бактерии с помощью жгутиков или ресничек. Жгутики имеют вид более или менее длинных нитей. Они очень нежны, тонки, легко обрываются, без специального окрашивания в микроскоп не видимы. Диаметр их не превышает 1/20 части поперечника тела бактерии.
Подвижные формы бактерий делятся на следующие группы:
- монотрихи - есть только один полярный жгутик,
- лофотрихи - имеется пучок жгутиков на одном из концов клетки,
- перитрихи-жгутики расположены по всей поверхности тела.
Характер расположения жгутиков на теле бактерии определяет и характер ее движения - прямолинейное или беспорядочное. Подвижность бактерий зависит от ряда условий: температуры, состава питательной среды, продуктов их жизнедеятельности и т. п. Подвижные формы бактерий не во всех стадиях своего развития и не во всех условиях роста снабжены жгутиками.
Спорообразование
В теле многих бактерий в известные периоды их развития появляются круглые или эллипсоидальные образования - опоры. Они обычно завершают цикл развития бактерий. Величина спор по сравнению с величиной произведших их клеток может колебаться в широких пределах.
Опоры образуются не у всех видов бактерий. Они окружены хорошо обособленной оболочкой, почти непроницаемы для воды и являются наиболее устойчивыми образованиями среди всего живого мира. Так, они нередко выдерживают кипячение в течение нескольких часов и продолжительное действие сухого пара при температуре от 120 до 140°. Споры многих бацилл сохраняют свою жизнеспособность после продолжительного пребывания при температуре -190° и даже при температуре жидкого водорода (-253°). Устойчивы они также и к действию химических веществ - ядов. Все это чрезвычайно затрудняет борьбу с болезнетворными споровыми видами бактерий.
Зрелая спора в течение десятков лет может сохранять свою жизнеспособность. Обычно прорастание спор происходит после некоторого периода покоя и связано с воздействием внешних условий. Весь процесс спорообразования протекает в течение суток и менее. После созревания споры, произведшая ее клетка постепенно отмирает и зрелая спора выходит наружу. При прорастании она набухает, становится богаче водой и из нее выходит проросток, одетый в тонкую оболочку.
Размножение бактерий
Достигнув состояния зрелости и предела роста, бактерии начинают размножаться простым делением. При делении в средней части тела бактерии появляется перегородка, которая затем расщепляется и обособляет две новые клетки. Последовательное расположение перегородок при делении у разных бактерий различно. У палочковидных форм оно располагается перпендикулярно к длинной оси, у шарообразных форм перегородки могут располагаться в одной, двух или трех взаимно-перпендикулярных плоскостях, с чем и связано образование таких форм, как стрептококки, тетракокки и сарцины.
Быстрота размножения бактерий зависит от ряда условий и может быть весьма различной. Чем благоприятнее внешние условия существования бактерий, тем быстрее происходит их деление. В нормальных условиях число бактерий удваивается приблизительно через каждые полчаса. Вели бы оно всегда происходило беспрепятственно, то количество бактерий от одной клетки достигло бы колоссальных размеров. По подсчету микробиолога Кона, потомство одной бациллы через пять дней могло бы заполнить все моря и океаны. Однако этого никогда не было и никогда не будет. Жизненный цикл бактерий ограничен определенными внешними условиями, за пределами которых размножение замедляется или вовсе приостанавливается. Недостаток питания, вредные продукты обмена, конкуренция различных видов и т. п. губительно действуют на бактерии. При неблагоприятных условиях они массами погибают.
Классификация бактерий
Положение бактерий в системе живых существ пока еще недостаточно определено. Принято считать, что бактерии представляют часть растительного мира, а грибы и водоросли являются ближайшими к ним родственными организмами. Морфологические признаки бактерий в большинстве случаев ограничены немногими формами: шаровидны, палочки, спирали. Необычайная простота и элементарность их внешней организации затрудняют их классификацию. Определение вида бактерии на основе только морфологических признаков невозможно. Научная систематика основана на морфологии и истории развития, но для классификации бактерий необходимо знать не только морфологию, но и их физиологические и биохимические особенности. В связи с этим устанавливаются: отношение бактерий к кислороду, температурным условиям, образование пигмента, разжижение желатина, образование кислот и газа на сахарах, изменение молока при росте в нем бактерий, образование индола, сероводорода, аммиака, редукция нитратов в нитриты или в свободный азот. Однако и этого не всегда бывает достаточно для определения вида бактерии.
Существуют различные системы классификации бактерий, но все они являются условными, и далекими в большей или меньшей мере от естественной классификации. Рассмотрение этих систем или хотя бы одной из них в данном случае не является необходимым даже в применении к фитопатогенным бактериям. Следует лишь сказать, что в настоящее время почти все фитопатогенные бактерии объединены в родах Pseudomonas, Xanthomonas, Bacterium и Erwinia.
В последнее время М. В. Горленко (1961) предложил следующую систему классификации фитопатогенных бактерий класса Eubacteriales:
I. Семейство Mycobacteriaceae (Честер, 1901)-неподвижные бактерии (без жгутиков):
- 1-й род - Gorynebacterium (Леман и Нейман, 1896) - (грамположительные бактерии;
- 2-й род Aplanobacterium (Смит, 1905, Гешич, 1956) - грамотрицательные бактерии.
II. Семейство Pseudomonadaceae (Вильсон и др., 1917) - бактерии со жгутиками (полярными):
- 1-й род - Pseudomonas (Мигула, 1900) - бактерии неокрашенные и флюоресцирующие;
- 2-й род - Xanthomonas (Доусон, 1839) - бактерии с окрашенными колониями.
III. Семейство Bacteriaceae (Кон, 1872) - подвижные бактерии с перитрихальными жгутиками, не образующие опор:
- 1-й род - Bacterium (Эренбергер, 1828) - неокрашенные формы, не образующие пектиназы и протопектиназы;
- 2-й род - Pectobacterium (Уолди, 1945) - неокрашенные формы, образующие пектиназу и протопектиназу;
- 3-й род - Chromobacterium (Бергонцини, 1881) - окрашенные формы.
IV. Семейство Bacillaceae (Фишер, 1895) - подвижные бактерии, спорообразующие палочки:
- 1-й род - Bacillus (Кон, 1832) - клетки при спорообразовании не вздуваются или вздуваются слабо;
- 2-й род - Clostridium (Празновский, 1880) - клетки при спорообразовании вздуваются.
В приведенной выше системе общепринятый до сих пор род Erwinia опущен. Из него выделяется особый род Pectobacterium, в который включаются бактерии с перитрихальными жгутиками и обладающие пектолитической активностью. Те из фитопатогенных бактерий, которые такой способностью не обладают, отнесены к роду Bacterium. Эта система, рациональная сама по себе, нова и пока еще не вошла в быт, поэтому в настоящей работе мы придерживаемся той классификации, в которой роду Erwinia отводится свое место. Такое родовое название фитопатогенных бактерий широко применяется в специальной литературе как в нашей стране, так и за рубежом.
Определение вида бактерий без применения искусственных питательных сред невозможно. В связи с этим можно отметить, что при культивировании бактерий образуют весьма характерные колонии. В таком случае по одному внешнему виду можно судить о видовой принадлежности бактерий.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
1.5. Современная классификация бактерий В современной систематике бактерий сложилась ситуация, характер- ная и для классификации других организмов: достигнуты успехи в соз- дании филогенетической системы классификации, отражающей основ- ные направления эволюционного развития и родство представителей оп- ределенных таксонов, но сохраняют свое значение искусственные фено- типические классификации, более удобные для идентификации микроор- ганизмов. В настоящее время отсутствует сколько-нибудь детализированная эволюционная система прокариот и, скорее всего, решение этой пробле- мы – дело неблизкого будущего. Особенности прокариот в области мор- фологической, физиолого-биохимической, генетической организации го- ворят о неприменимости к ним хорошо разработанных принципов, ис- пользуемых при построении системы высших организмов. Не останавливаясь на исторических аспектах проблемы систематики бактерий, следует отметить, что наиболее приемлемой филогенетической системой классификации прокариот является система, основанная на со- поставлении последовательности нуклеотидов в 16S-рРНК. Эта система положена в основу 2-го издания многотомной энциклопедии прокариот – Bergey’s Manual of Systematic Bacteriology (Руководство по систематике бактерий Берджи), первый том которой вышел в свет в 2001 г. В этом труде все прокариоты разделены на 26 филогенетических «ветвей» (групп) на основании строения их 16S-рРНК; 23 «ветви» представлены эубактериями, а три – архебактериями. Следует подчеркнуть, что боль- шое количество этих филогенетических групп содержат виды прокариот, которые не выделены в виде чистых культур и поэтому еще детально не изучены. Для представителей данных видов известны в настоящее время только последовательности нуклеотидов в 16S-рРНК. Из 23 групп эубак- терий две филогенетические группы представлены грамположительными бактериями, остальные группы – грамотрицательными. Грамотрицательные бактерии состоят из крупной группы Протеобак- терий (Proteobacteria) и 20 групп остальных бактерий, имеющих данный тип клеточной стенки. Краткая характеристика Протеобактерий, к кото- рым по составу 16S-рРНК наиболее близки митохондрии и хлоропласты большинства эукариот, приведена в табл. 2. Протеобактерии – очень гетерогенная в морфологическом, физиоло- гическом и биохимическом плане группа грамотрицательных бактерий. Для представителей этой группы характерны все типы энергетического метаболизма и питания. Клетки большинства видов Протеобактерий имеют палочковидную, сферическую или вибриоидную форму, размно- жаются в основном бинарным делением, но для некоторых видов харак- терно почкование и образование плодовых тел в сложном клеточном цикле. В этой группе имеются как подвижные за счет жгутиков, так и неподвижные бактерии. По отношению к молекулярному кислороду Протеобактерии бывают облигатными аэробами, облигатными и факуль- тативными анаэробами. Группа Протеобактерий на основании различий в 16S-рРНК разделена на пять подгрупп: альфа, бета, гамма, дельта и эп- силон. Кроме Протеобактерий, к грамотрицательным относятся следующие основные группы эубактерий: водородные термофилы, зеленые нитчатые бактерии, зеленые серные бактерии, цианобактерии, спирохеты, цитофа- ги, бактероиды, хламидии, планктомицеты, дейнококки, хлорофлексусы, фузобактерии, фибробактерии, термодесульфобактерии и др. Филогенетические группы грамположительных бактерий – Actinobacteria и Firmicutes. Группа Actinobacteria («актиномицетная ветвь») пред- ставлена следующими родами бактерий, имеющими в ДНК высокое со- держание ГЦ-пар: Geodermatophilus, Frankia, Streptomyces, Arthrobacter, Micrococcus, Actinomyces, Bifidobacterium, Propionibacterium, Actinoplanes, Nocardia, Rhodococcus, Corynebacterium, Mycobacterium. Группа Firmicutes («клостридиальная ветвь» – главным образом грамположи- тельные бактерии с низким содержанием ГЦ-пар в ДНК) состоит из следующих родов: Clostridium, Lactococcus, Pediococcus, Streptococcus, Enterococcus, Leuconostoc, Listeria, Caryophanon, Staphylococcus, Sarcina, Sporosаrcina, Bacillus, Desulfotomaculum, Heliobacterium, Mycoplasma,Ureaplasma и др. В составе архебактерий выделяют три филогенетические группы: Crenarchaeota, Euryarchaeota и Korarchaeota. Группа Crenarchaeota со- стоит из экстремально термофильных бактерий, большинство представи- телей которых осуществляют метаболизм серы, некоторые восстанавли- вают ионы железа и молибдена. В группу Euryarchaeota входят облигат- но анаэробные метаногенные архебактерии, а также экстремальные тер- мофилы и галофилы. Группа Korarchaeota образована архебактериями, обитающими в горячих серных источниках. До настоящего времени ни один из представителей этой группы (обладающих сходной 16S-рРНК) не выделен в виде чистой культуры, поэтому их фенотипические призна- ки изучены недостаточно. Заканчивая рассмотрение филогенетических ветвей прокариот, сле- дует отметить, что предложенная филогенетическая система, основанная на исследовании нуклеотидных последовательностей только одного гена рибосомной РНК – не более чем одна из технически удобных и разрабо- танных систем упорядочения многочисленных организмов в целях их идентификации, поэтому построить логически верную таксономию бак- терий только с учетом этого признака не представляется возможным. Наиболее признанной и используемой фенотипической классифика- цией бактерий является классификация, представленная в девятом изда- нии Определителя бактерий Берджи. В этом издании бактерии на осно- вании строения пограничного слоя клетки разделены на четыре основ- ные категории (отдела): 1) Gracilicutes (от лат. cutes – кожа, gracilis – тонкий) – грамотрицательные эубактерии, имеющие клеточные стенки; 2) Firmicutes (от лат. firmus – прочный) – грамположительные эубакте- рии, имеющие клеточные стенки; 3) Tenericutes (от лат. tener – мягкий, нежный) – эубактерии, лишенные клеточных стенок; 4) Mendosicutes (от лат. mendosus – ошибочный) – архебактерии, клеточные стенки которых отличаются от аналогичных структур других прокариот. В отдел Gracilicutes входят бактерии различной морфологии с гра- мотрицательной клеточной стенкой. Размножение происходит в основ- ном бинарным делением, некоторые бактерии размножаются почковани- ем. Эндоспор не образуют. Большинство подвижны: встречаются все ти- пы передвижения бактерий – с помощью жгутиков, скольжением, изги- банием. Отдел включает аэробные, анаэробные и факультативные ана- эробные бактерии; фототрофные и хемотрофные бактерии. Отдел под- разделяют на три класса: Scotobacteria, Oxyphotobacteria, Anoxyphotobacteria. В класс Scotobacteria входят грамотрицательные бактерии, не ис- пользующие световую энергию для целей метаболизма, а получающие ее только в результате окислительно-восстановительных реакций. Название класса происходит от греч. sсotos – темнота. Это самый крупный класс бактерий. В класс Anoxyphotobacteria входят пурпурные бактерии, зеле- ные бактерии и гелиобактерии, осуществляющие аноксигенный фото- синтез (без выделения молекулярного кислорода). Класс Oxyphotobacteria представлен цианобактериями и прохлорофитами, осуществляющими оксигенный фотосинтез (с выделением молекулярного кислорода). Этот тип фотосинтеза аналогичен фотосинтезу, протекающему в растениях. В отдел Firmicutes включены бактерии с грамположительной кле- точной стенкой. Клетки могут иметь разную форму: палочки, кокки, ни- тевидные, ветвящиеся. Некоторые представители образуют эндоспоры. Большинство из них неподвижны; подвижные формы имеют перитрихи- альное жгутикование. В состав отдела входят аэробные, анаэробные и факультативно анаэробные бактерии. Отдел состоит из двух классов: Firmibacteria, Thallobacteria. Класс Firmibacteria включает большое ко- личество «неветвящихся» грамположительных бактерий. Класс Thallobacteria включает бактерии, клетки которых способны «ветвиться». Отдел Tenericutes представлен бактериями, не имеющими клеточной стенки. В связи с отсутствием клеточной стенки форма клеток непосто- янна: в чистой культуре одного вида одновременно присутствуют кокко- видные, палочковидные, нитевидные, грушевидные, дисковидные и дру- гие клетки. Размножение бактерий, входящих в этот отдел, происходит бинарным делением, почкованием. Окрашивание по Граму отрицатель- ное. Характерно образование мелких, врастающих в агар колоний. Могут быть сапрофитными, паразитами или патогенами. Отдел состоит из од- ного класса Mollicutes (микоплазмы). Отдел Mendosicutes образован бактериями с ригидной клеточной стенкой, но не содержащей пептидогликана муреина. Большинство пред- ставителей – строгие анаэробы, многие из которых имеют жгутики. Ви- ды характеризуются экологическим и метаболическим разнообразием, способностью жить в экстремальных условиях. Отдел состоит из одного класса – Archaebacteria. В составе четырех отделов (основных категорий) выделено 35 групп (или секций) бактерий, которые в большей или меньшей степени будут охарактеризованы в последующих главах. К отделу Gracilicutes принадлежат следующие группы. Группа 1. Спирохеты. Группа 2. Аэробные (или микроаэрофильные), подвижные, спирале- видные (или вибриоидные) грамотрицательные бактерии. Группа 3. Неподвижные или редко подвижные грамотрицательные изогнутые бактерии. Группа 4. Грамотрицательные аэробные (или микроаэрофильные) па- лочки и кокки. Группа 5. Факультативно аэробные грамотрицательные палочки. Группа 6. Грамотрицательные анаэробные прямые, изогнутые или спиралевидные палочки. Группа 7. Бактерии, осуществляющие диссимиляционное восстанов- ление серы или сульфата. Группа 8. Анаэробные грамотрицательные кокки. Группа 9. Риккетсии и хламидии. Группа 10. Аноксигенные фототрофные бактерии. Группа 11. Оксигенные фототрофные бактерии. Группа 12. Аэробные хемолитотрофные бактерии и близкие организ- мы. Группа 13. Почкующиеся и (или) образующие выросты бактерии. Группа 14. Бактерии, имеющие чехлы. Группа 15. Нефотосинтезирующие скользящие бактерии, не обра- зующие плодовых тел. Группа 16. Скользящие бактерии, образующие плодовые тела. В отдел Firmicutes входят: Группа 17. Грамположительные кокки. Группа 18. Грамположительные палочки и кокки, образующие эндо- споры. Группа 19. Грамположительные палочки правильной формы, не обра- зующие спор. Группа 20. Грамположительные палочки неправильной формы, не об- разующие спор. Группа 21. Микобактерии. Группы 22–29. Актиномицеты. К отделу Tenericutes принадлежит: Группа 30. Микоплазмы. Отдел Mendosicutes включает: Группа 31. Метаногены. Группа 32. Сульфатредуцирующие архебактерии. Группа 33. Экстремально галофильные архебактерии (галобактерии). Группа 34. Архебактерии, лишенные клеточной стенки. Группа 35. Экстремально термофильные и гипертермофильные архе- бактерии, метаболизирующие серу. В заключение следует подчеркнуть, что большинство микроорганиз- мов, существующих в природных сообществах, еще должно быть выде- лено в чистые культуры. Считается, что в настоящее время культивиро- вать можно только 0,1 % всего микробного разнообразия, а остальных представителей бактерий вырастить и идентифицировать не удается, хо- тя уже в чистую культуру выделены и описаны около 5 тыс. видов про- кариот.